(I recommend the reader consult the "preface" or "guide" to this paper, which may be found at "About the Papers: An Introduction" (section III)).
Abstract
(See: The
Fractal Organization of Nature (table)) (also at the
bottom of this paper)
A General Systems model of the Universe is presented, based on a fractal paradigm. A 4x3 "metric" or fractal algorithm is proposed as the organizing principle of the Microphysical, Biophysical, Astrophysical, and Metaphysical realms. The algorithm is expressed through the conservation laws and forces of physics, symmetry-breaking and the creation of matter during the "Big Bang", the dimensions of spacetime, the quantum rules of atomic order, the families of elementary particles, the structure of baryons, alpha particles, carbon atoms, carbon's molecular and crystal bonds, DNA, the genetic code, the nucleosynthetic pathway of stars, gravitation, etc. The human mind has discovered this order both intuitively and rationally, where it finds expression in the 4x3 (and 4x4) patterns of mythological, occult, religious, philosophical, and scientific "world systems" and cosmologies.A model of the unified field theory is presented as an example of how the fractal algorithm may be used in system modeling. In its most general form, the 4-part algorithm consists of an input of energy, followed by two complementary conservation modes, then concluding with a restorative force which either returns the system to its actual original state, or elevates it to a "harmonic complexion" or resonant analog of the original state.
The concept of hierarchical order is fundamental to General Systems theory (Bertalanffy, 1968). The exposition of fractal geometry (Mandelbrot 1983) raised a new paradigm of natural order which offered a theoretical and mathematical basis for systems generally and their hierarchical characteristics specifically: it is the fractal nature of the Universe we see in its hierarchical structure, systemic character, and organic connectivity. Self-similar replication and harmonic resonance are natural features of fractal structure and organization.
Physical evidence for the operation of a universal 4x3 fractal algorithm producing a resonant cosmic order is presented in table 1. This is a hierarchical arrangement which encompasses 4 realms: Microphysical, Biophysical, Astrophysical, and Metaphysical (human conceptual). The fractal algorithm which produces this hierarchy is a 4x3 complex drive created by free energy (light) in three conservation modes: Entropy (dimensionality, intrinsic motion, and gravitation), Raw Energy (matter and momentum), and Symmetry (charge, spin, inertia and gravitation (again)). The emergent properties of this system in operation often require a 4x4 representation. (See: ""Unified Field Table Simple Form".)
Each realm of the hierarchy is characterized by a conspicuous center of mass, energy, or information, the site of the 4x3 fractal algorithm or energy pattern which organizes the general structure of the realm. In the Microphysical Realm this center is the atomic nucleus; in the Biophysical Realm it is the nucleus of the cell; in the Astrophysical Realm it is the galactic nucleus; in the Metaphysical Realm, it is the human brain, contemplating body/spirit (intuitive mode), or unified theories of matter/energy (rational mode). Individual levels within each realm have their own centers of structure, dynamics, and/or information.
Such a universal pattern can have only the most general of characteristics. I have designated its horizontal 4-component in structural terms as: Unit, Pair, Group, Emergent new unit; the group category is the locus of the 4x3 energy pattern which produces the Emergent new unit. This new unit then becomes the basis of the next level in the hierarchy. George Land (1973) and August Jaccaci (1980) have characterized this structural series in dynamic terms as: Accretive, Replicative, Mutualistic (Land), or: Gather, Repeat, Share, Transform (Jaccaci). The vertical 3-component of the matrix (plus its 4th emergent level) Jaccaci identifies as a simple repetition of the horizontal series. In its most complete form ("World Systems"), these parameters resolve to: 1) an input of free energy followed by symmetry-breaking, establishing a joint dimensional conservation domain (spacetime) for light and matter; 2, 3) two complementary conservation modes: 2) raw energy conservation including entropy (mass, momentum, kinetic and potential energy, and low-energy gravitation); 3) symmetry conservation (charge, spin, inertia, intrinsic motion, and high-energy gravitation); 4) restorative, transformative, or evolutionary forces (the 4 forces of physics) which either return the system to its original state immediately (as in matter-antimatter annihilations), or through time ("quantum radiance" of black holes, proton decay), or evolve a resonant analog of the original state (the various material levels of the hierarchy - entropy, symmetry, and gravitationally driven "harmonic complexification").
Readers familiar with the theory of "Chaos" may recognize a familiar pattern in the model: beginning with a "linear" or simple input (cell 1), passing to a stage of bifurcation (cell 2), progressing to a stage of greater complexity including secondary bifurcations (cell 3), and finally a "chaotic" stage (cell 4) which predictably settles in the "fractal basin" of a 4x3 (or 4x4) "strange attractor" (the precise physical nature of the attractor, however, is unpredictable). Like a true fractal algorithm, the output of one level becomes the input of the next, the self-feeding iterations producing new self-similar expressions at ever-higher levels of information and system complexity. The model is scale-invariant, not only vertically with each row, but also with respect to the four major realms. (See: "Chaos: Making a New Science" by James Gleick, Viking Penguin Inc., 1987)
(See also: "The Fractal Organization of Nature"; "The Information Pathway"; "Teilhard de Chardin: Prophet of the Information Age").
The Microphysical Realm
(atomic matter, nuclear organization, and chemistry: 3 particles
(protons, neutrons, electrons) x 4 quantum numbers (n,l,m,s)
(See: "Table No. 1")
Level 1: Elementary
Particles
(3 energy "families" each of 4 elementary particles)
We begin the hierarchy with a defining event, the production of elementary particles in the "Big Bang" (row 1). While we know the products of this first level, we do not really know how to initiate it. What do we put in the first cell of row 1 as the primordial unit of energy and organization from which all else evolves? We can of course simply put the Big Bang or First Cause in this first cell, claim Divine Intervention, confess our ignorance, and leave it at that. Personally, I am not fundamentally opposed to this solution; we will never know everything and a frank admission of this fact from the outset is intellectually healthy and honest. Nevertheless, in the light of present knowledge, it is possible to fill out the first horizontal row of the hierarchy in sensible, physical terms without presuming that the physical reality is the only reality, that we have done anything more than solve a mechanical puzzle, or that we know, or can know, anything at all about the fundamental origins of free energy (light) itself. (See also: "The Higgs Boson and the Weak Force IVBs", to extend the hierarchy (or the details of the first cell) to even higher energies and earlier times.)
With these apologies, in the first cell of Level One, I place high-energy light - free electromagnetic radiation - the most symmetric and therefore the most primitive form of energy we know. In the second cell of row one, which is the position of the pairing function, I place space-time, the conservation domain and metric of light, created by light's own intrinsic motion or entropy drive, gauged by "velocity c". Along with spacetime in the second cell are several other possible pairings: matter-antimatter and the particle-wave duality of light. We can view spacetime as the symmetric product of light's wave form, and the matter-antimatter field as the symmetric product of light's particle form. So long as the matter-antimatter field remains symmetric, it will appear as light only; once its symmetry is broken, however, it will manifest as particles. Dualities are fundamental throughout nature; other dualities exhibited by light include its oscillating electric vs magnetic energy fields, and the "frequency vs wavelength" characterization of its physical wave structure. (Frequency x wavelength = c, the electromagnetic constant or the "velocity of light".)
Spacetime is regulated by a combination of energy and symmetry-conserving "inertial" forces, and by a number of "gauge" constants which regulate various conservation parameters (c, G, e, h, etc.). Spacetime is actually created by the entropy drive of free energy, manifesting as the intrinsic motion of light. Spacetime is the entropic conservation domain of light, and it contains a "metric" or structural order which is the enforcer of spacetime's conservation role (the source of "inertial" forces). This metric determines or "gauges" the value of c, the velocity of light, and by this means fixes the measured and energetic equivalence between each of the four dimensions. (Without the metric, every photon could be a "law unto itself" with a unique "personal" velocity.) Hence light and spacetime themselves, far from being simply an amorphous waveform of energy, possess an internal structural order from the beginning, an order required by energy conservation. This implicit order also has an alternative form, well defined, regulated, and also symmetric, the matter-antimatter particle pairs which populate the "virtual particle sea" of the "vacuum". These particle pairs occur in a relatively small number of quantized forms belonging to the quark and lepton families. The virtual particles (cell 3) become real when the weak force breaks the symmetry of matter-antimatter particle pairs during the initial moments of the "Big Bang" (cell 4).
The virtual particle "zoo" is here interpreted as the natural product of the interaction of light's energy with its own structural spacetime metric. Thus the "particle metric" bears a structural imprint from the spacetime metric. In the "standard model" of particle physics, the "Higgs boson" or "Higgs field" is the presumed source (regulator) of elementary particle mass. In the (similar) view presented here, the spacetime metric determines the possible forms that particles may take, while the Higgs boson determines the energy level (environment) in which "singlet" elementary particle creation and/or transformation may occur. (See: The "Higgs" Boson and the Weak Force IVBs: Part I for a detailed explanation.)
In the the third cell of row 1, which is the "group" cell of interaction (Jaccaci's "sharing of differences", Land's "mutualistic" cell) we can place for simplicity of representation a virtual matter-antimatter "leptoquark" pair, which stands for the interaction between the mass-bearing quark field of the strong force "color" charge, and the charge-bearing leptonic field of the weak force "identity" charge. In this third cell we usually find an expression of Nature's 4x3 fractal algorithm; in this first row we find three families of four particles (six leptons and six quarks), which decay (in cell 4) to their ground states as proton, electron, and electron neutrino (a leptoquark antineutrino is presumably also produced to balance the proton's "baryon number charge").
In cell four, interactions among and between the quarks and leptons are mediated by the weak force "Intermediate Vector Bosons" (IVBs), the W+, W-, and Z, while leptoquark decay is mediated by a similar (but hypothetical) much heavier "X" boson (see "The Particle Table"). The final product of the asymmetric leptoquark decay is a particle of matter, the humble baryon, proton, or nucleus of the hydrogen atom. This baryon, which is literally the foundation stone of the material Universe, is also a 4x3 expression of the universal fractal algorithm: 3 quarks bearing 4 charges (charges relevant to the microphysical realm, excluding gravity) - electric, color, flavor, and spin. The first horizontal row of the hierarchy essentially recapitulates the reaction equation for the formation of matter in the "Big Bang". (See: "The Origin of Matter and Information".)
In the first line we have produced from the order and energy inherent in spacetime and light, the order and energy of the particulate realm of matter, passing from an initial symmetric state of free electromagnetic energy, which contained only potential information and metric structure, to a final asymmetric state of bound electromagnetic energy, which realizes that information and expresses that structure in manifest, particulate, conserved, and ordered form: the baryons and their alternative charge carriers, the leptons. While we do not understand the details of this process, what we do understand is that the energy, order, and information content of light and spacetime has been transformed, extracted, or projected from a symmetric potential to an asymmetric reality. The mass, order, and information content of particles does not come from nowhere; it comes from the metric of spacetime and the energy of light. (Note: leptoquarks and the X boson (and the "Higgs") remain beyond the energy range of our most powerful accelerators, and so are all still hypothetical. I include them because they (or their analogs) should exist if we are to make sense of the other confirmed elements of the theory).
The baryon, whose ground state is the proton, is the essential material, massive, and asymmetric product of the Big Bang. Baryons are composed of three quarks, the mass carriers of matter. The existence of matter depends upon the existence and stability of the baryon. The baryon, with its 3 quarks, 4 charges, and 8 interacting gluons (each composed of a color-anticolor charge), exchanging color charges between quarks at the speed of light, is a particle of incredible internal complexity. Within its invariant, beating heart of "sticky light" lies the mystery of bound electromagnetic energy (matter), and upon its stability rests every higher structure of the manifest Universe.
Matter is thought to be produced by the asymmetric weak force decay of electrically neutral leptoquark-antileptoquark pairs, a decay possibly mediated by the (hypothetical) "X" IVB. Electrically charged particle-antiparticle pairs cannot decay asymmetrically - they can only annihilate each other. Because matter can only be produced from the asymmetric decay of an electrically neutral particle, leptoquarks and baryons must be composite particles whose internal components can sum to electric neutrality (like a neutron). This is the fundamental reason for the existence of the sub-elementary quarks and their fractional electric charges. (See: "The Formation of Matter and the Origin of Information".)
By a similar logic, matter could also not exist without the essential service provided to baryons by the leptonic field of alternative charge carriers - massive leptons to carry electric charges (the electron and its kin), massless (or nearly massless) neutrinos to carry identity charges (the electron neutrino and its kin, including the leptoquark neutrino) (mesons also function as alternative charge carriers of quark fractional charges - flavor, color, spin, and electric charge - facilitating baryon transformations). (See: "The "W" IVB and the Weak Force Mechanism".) Without the leptonic field of alternative charge carriers, the quark field would remain trapped as symmetric, mutually annihilating, particle-antiparticle pairs. This is why matter consists of two fields, the sub-elementary mass-carrying quarks, or composite hadrons, and the alternative charge-carrying elementary particles, or leptons (see: "The Weak Force: Identity or Number Charge"). The (hypothetical) leptoquark, which is essentially an elementary lepton fractured internally into three parts, is the ancestor of both hadrons and leptons, which is why these two fields interact so seamlessly. Finally, there remains an asymmetry of unknown origin in the interaction of the weak force IVBs with matter vs antimatter, such that, for example, neutral antileptoquarks decay at a slightly faster rate than neutral leptoquarks, producing the residue of matter which composes our material Universe (a tiny asymmetry of (approximately) one part per ten billion).
Light and particles (free and bound forms of electromagnetic energy) are a thoroughly integrated duality in which the mass energy of particles is derived from the free energy of light (E = mcc), while the information content of particles is derived from the information implicit in the metric structure and conservation parameters of spacetime. Since light creates spacetime (via light's intrinsic motion or spatial entropy drive) as well as particle mass (via light's energy content) we see that matter is in many ways simply an alternative form of light. This is also the message of Heisenberg/Dirac "virtual particles", endlessly forming, annihilating, and reforming in the "vacuum" of spacetime. We therefore can expect to find echoes of spacetime and light in the realm of matter. Such were discovered in Einstein's famous relations between matter, energy, gravity, and spacetime (the Special and General Theories of Relativity), and extend to other phenomena as well: DeBroglie mass waves, to take an example from the quantum realm. Further examples of this duality include: local massive vs non-local massless energy forms; a conserved, historical entropic domain for bound energy, produced by the intrinsic motion of time, which is the analog of the conserved, spatial entropic domain for free energy produced by the intrinsic motion of light; negative, contractile forms of gravitational energy and entropy vs positive, expansive forms of light energy and entropy; the invariance of velocity c vs the invariance of charge. Symmetry parameters transformed from light to matter can be seen in the conservation terms of "Noether's Theorem": the charges of matter are the symmetry debts of light. This intimate union between light and matter is the basis of a Unified Field Theory as presented on my website. (See: "A Short Course in the Unified Field Theory", and is more properly and fully discussed there.
The great message from the atomic realm is that of order - a message intuitively recognized from ancient times in crystals. Life, the most complex expression of matter, could have arisen spontaneously only from such an orderly substrate. More recently, beginning with Planck and Einstein, we have realized that the order in crystals and the atomic realm, including quantum order, has its origin in the order inherent in light and spacetime - specifically in the four conservation laws of energy, entropy, symmetry, and causality (see: "The Tetrahedron Model". The atomic realm of matter and time is but an alternative, secondary expression of the metric realm of light and space. Both realms are strictly conserved, and transformations between them are strictly regulated. As we progress through the levels of the natural hierarchy below, remember that the order we are seeing in these material realms has its origin in light.
(See: "The Higgs Cascade and the Weak Force IVBs", an expansion of row 1 into a 4x4 General Systems model of the "Big Bang".) (See also: "The Origin of Matter and Information".)
Level Two: Atoms, Elements;
Nuclear Fission/Fusion
(the alpha particle: 4 nucleons each of 3 quarks)
The Atomic Nucleus and its Electron Shell
The baryon (proton) is the principle product of the "Big Bang" and the first rung of the structural hierarchy ladder. It is the fundamental unit of step two and is the foundation stone upon which is built the entire realm of matter. Baryons are the class of particles which contain 3 quarks, of which the proton and neutron are familiar examples. The baryon is an absolute marvel of construction, consisting of three quarks bearing "color" charges which are exchanged via an internal field of 8 gluons. Gluons carry color-anticolor charges (in any combination), are massless, and move at velocity C. Gluons have been compared to "sticky light". The "round-robin" exchange of color charges between quarks constitutes the strong force, which keeps quarks permanently confined in baryons and whose conserved presence also prevents "proton decay", the complete disintegration of a baryon via a leptonic (weak force) decay, possibly mediated by the (hypothetical) "X" IVB.
Quarks occur in six species, or "flavors", of which the u (up) and d (down) quarks are the "ground" or lowest energy states. The quark composition of the positively charged proton is (uud)+, that of the electrically neutral neutron (udd). These are the familiar "nucleons". Exotic baryons containing heavier flavors are known as "hyperons". (See: "The Particle Table".) Quark flavors can transform one into another if there is sufficient energy, and if a leptonic (and/or meson) alternative charge carrier can be produced by the "W" IVB of the weak force. Baryons carry a lot of information with their three quarks, two flavors, various spin orientations, multiple color and electric charges, and their internal gluon field. This is a highly energetic, highly organized, permanently bound, extremely stable, mass unit of high internal information content - and it will interact with others of its own kind to produce extremely complex compound nuclei (in Nature, up to 238 nucleons in the uranium atom). This is indeed a very promising foundation particle upon which to build the information pathway.
In the second cell of row 2, for the pairing function of the baryon we have the up, down (u, d) quark pair, and all the gluon pairs; meson pairs belong at this level as well. (Gluons are composed of color-anticolor charges, and carry color charges between quarks. Mesons are composed of quark-antiquark pairs, and (among other services) carry fractional flavor, electric, spin, and color charges between quarks, especially in the weak force transformations of baryons). At the level of the whole atom, we have the electron-proton charge pair; within the electron shell, we have the north-south magnetic pairings of the electrons in their shared orbits (spin up/down). The particle level is particularly rich in pairings; we could name more. Perhaps the most important of these many pairings is that between the leptons (alternative charge carriers) and hadrons (mass carriers), illustrated here by the electron and proton.
In the proton-electron pair, or hydrogen atom, the negative electrical charge of the electron balances the positive electrical charge of the proton. The electron of row 2 is performing the same electrical function for the proton as the antiparticle of row one performed for matter, but of course the electron is not the antiparticle of the proton. This difference emphasizes the essential asymmetry of matter and the role of leptons as alternative charge carriers for the baryons - alternative in that they are not antiparticles but nevertheless carry (some of) the charges of antiparticles. The function of alternative charge carriers (which include mesons and the nearly massless neutrinos as well as the massive leptons) is crucial in that they allow the possibility of charge balancing and conservation without the annihilation reactions of antiparticles. Without the leptonic alternative charge carriers, the quark mass field could never break the symmetry of matter-antimatter leptoquark pairs; without the meson field, baryons, once formed, could not be transformed. (See: "The Origin of Matter and Information"); (See: "The 'W' IVB and the Weak Force Mechanism".)
At the level of the compound nucleus we find the proton-neutron pair; the exchange of virtual mesons between protons and neutrons (producing "nucleons") is responsible for the strong force binding of compound atomic nuclei. The binding of nucleons within heavy nuclei is another pairing function, an expression of the "Yukawa" strong force involving "flavor" charge exchange between baryons (via meson field vectors), rather than the "Gell-Mann" strong force involving "color" charge exchange between quarks (via gluon field vectors). (See: "The Strong Force: Two Expressions".)
In the third cell of row 2 (the "group" function) we have the helium nucleus (commonly referred to as an "alpha" particle), the product of the nuclear fusion of 2 protons and 2 neutrons. Helium is the second element of the periodic table, and its nucleus or alpha particle, because of its extreme stability, becomes the preferred building block or "brick" of the nucleosynthetic pathway in stars. This pathway, with the aid of supernova explosions, builds the remainder of the 92 elements of the periodic table. Because the alpha particle "brick" is the energetically favored building block of nucleosynthesis, elements with even atomic numbers (numbers of protons) are typically more abundant than elements with odd atomic numbers.
Carbon is the second major product of the nucleosynthetic pathway after helium. Both helium and carbon are perfect nuclear resonances of the 4x3 fractal pattern. Helium is composed of 4 nucleons each of 3 quarks; carbon is composed of 3 alpha particles (helium nuclei), each of 4 nucleons. The carbon nucleus is therefore a triple 4x3 resonance (the baron itself is the first resonance), unique among the atoms of the periodic table. The electron shell of carbon contains yet another 4x3 resonance, with four valence electrons in its third shell, the only atom with both a nuclear and an electron shell 4x3 resonance. We will discuss the remarkable and portentous 4x3 carbon resonances further in the next two levels.
Protons and neutrons can join together in high-energy fusion reactions in the center of stars, initiating the nucleosynthetic pathway, building the compound nuclei of the heavy elements. As each proton is added to the nucleus, a compensatory electron is added to the electron shell, balancing the overall electric charge of the atom. The heavy, high-energy, stable and "conservative" nucleus holds the atom together, preserving its identity and integrity, while the light, low energy, changeable and "social" electron shell permits temporary interactions with other atoms. It is this special combination of nuclear stability with chemical flexibility that gives our macroscopic world both diversity and activity, while maintaining a certain degree of solidity and reliability.
The electron shell provides a low energy pathway for atoms to interact; nuclear interactions are at least a million times more energetic than those of the electron shell, with a correspondingly high energy barrier which must be overcome to initiate them (hence fusion occurs naturally only in the "Big Bang", supernovas, and the interiors of stars). It is a general rule throughout nature that reducing the energy barrier to interaction catalizes information building.
The 92 naturally occurring elements of the periodic table are the product of row 2. Of these, at least 25 are known to be required for biological life forms, and doubtless every one finds a use somewhere in Nature - including providing a suitable planetary, stellar, and galactic environment for life.
Level three - Molecules,
Chemistry, Organic Polymers, Crystals
(the carbon atom: 4 bonding sites in the 3rd electron shell)
Step three is the realm of chemistry, inorganic and organic; the formation of compounds, crystals, and polymers; and every type of interaction between the electron shells of atoms. The atom with its electron shell is the unit of the first cell of level 3.
The electron shell develops as a series of concentric "orbits" of complex geometry, and it is typically only the outer orbits which can interact with those of another atom. These interactions can take place only if there are vacant positions in the outer orbitals which can be filled by sharing electrons with another atom which also has vacant positions. This type of bonding is called "covalent", in reference to the shared orbital relationship, and provides a strong and stable bond, which is featured in the second cell of row 3. Other pairings at this level include the common atomic pairs, such as H2, O2, and N2, and the dipolar electric/magnetic character of chemical bonds generally.
We place the carbon atom in the third cell of row 3. Amazingly, in addition to carbon's triple nuclear 4x3 resonance pattern, it also contains a 4x3 resonance in its electron shell, having 4 bonding sites in the third shell (1S2, 2S2, 2P2). Carbon is the only atom in the periodic table with both a nuclear and electron shell 4x3 resonance. Carbon's molecular tetrahedral bonding structure creates yet another 4x3 resonance - a tetrahedron being composed of 4 triangles. Hence carbon contains five full 4x3 resonances, three in its nucleus, one in its electron shell, and one in its molecular bonding pattern (for example, diamond), a most remarkable and unique atom, with a most remarkable and unique potential.
In the fourth cell of row 3 we have crystals, and especially the organic polymers of carbon. It is of course the ability of the carbon atom, with its four bonding sites, to bond with itself in tetrahedra, chains, branching chains, rings, spirals, and many other forms of great length, molecular weight, and apparently infinite variety in their combinations, permutations, reflections, polymers and isomers, that sets the stage for the information explosion of "organic" or carbon chemistry, the molecular basis of the complex information systems of life. For example, the nuclear fusion of hydrogen with carbon can produce only the element nitrogen with an isotope or two; the chemical combinations between the electron shells of hydrogen and carbon, however, produces the immense class of the hydrocarbons, with billions of chemically distinct isomers. This is an especially striking example of how by lowering the energy level and barrier for interaction, the possibilities for information building are enormously magnified.
Level four - Cyclic,
Functional Chemical Systems; RNA-DNA
(RNA-DNA: 4 nucleotides x 3 functional groups)
In row 4 we enter the complex realm of functional, cyclic chemical systems, prebiotic chemistry, including the replicating molecule of RNA. Organic chemistry is characterized by covalent bonding ("hard bonds") between carbon atoms which build strong, stable chains or skeletal structures; these chains provide anchor points for functional chemical groups which interact with each other through weaker, ("soft") bonds. "Soft" bonds are more in the nature of attractive and repulsive electrical interactions between molecules rather than orbital bonds. "Soft" bonds include hydrogen bonds, ionic bonds, bonding due to Van der Waals forces, and hydrophilic/hydrophobic interactions. The geometry, shape, size, symmetry, and configuration of molecules is crucially important to their organic/biological properties, and these weak bonding forces are critically important to this level of information building. Again, reducing the energetic threshold of interaction is very important to the diversity of possible interactions, and hence also to building the information content of systems such interactions may produce. These "soft" and flexible molecular bonds, crucial to the complex forms of organic chemistry and biochemistry, we place in cell 2. Here too we place the base pairings of the 4 nucleotides of RNA (AU, CG) (adenine, uracil, cytosine, guanine).
Replication is the translation of the ability of carbon atoms to bond with their own kind (including the general phenomenon of crystal formation and growth), into the ability of carbon based macromolecules to bond with plus replicate their own kind. What produces crystal growth on one level produces replication on the next. It is a stage transition to a higher level of both organization and information, and the direct projection of nuclear, electron shell, and crystal bonding patterns into the realm of macro-molecular interaction. The pathway is directly through the 4x3 structure of carbon, which as we have seen, contains a 4x3 resonance in its nucleus, electron shell, and molecular bonding pattern. The covalent bonding of carbon with itself into stable branching chains and cyclic structures provides attachment points for functional groups involving other elements. These in turn can form "soft" hydrogen bonds with each other, producing macro-molecular chains of cyclic or periodic character, leading directly to the 4x3 resonant structure of RNA. Water itself, the "universal solvent", also produces a tetrahedral molecular bond; the combination of water and organic chemistry is literally pregnant with the potential for the 4x3 replicating molecular structures which will eventually lead to competitive, "self-interested" living forms.
In RNA, we again find a structural expression of the 4x3 fractal "metric". RNA consists of 4 nucleotides (adenine, guanine, uracil, cytosine) each composed of 3 functional groups - a nitrogenous base, a sugar, and a phosphate group (in DNA uracil is replaced by thymine). It is this 4x3 structure of RNA which is the centerpiece of row 4, cell 3.
Row four is the chemical threshold of life. Here we find carbon interacting with a variety of other elements to form the complex molecules which are capable of playing a role in future living systems. What is special about these molecules is that they do not simply link up with each other in inert crystalline combinations, they actually perform work of some sort, such as the capture and processing of energy, replication, the formation of defensive perimeters, etc. Why this sort of chemical activity should arise at all is a mystery: there is perhaps some advantage to be gained, on a level of molecular competition, by systems which can process daytime energy but also survive through the night. This survival function might include replication as a sort of energy-plus-memory storage.
The origin of life is perhaps more comprehensible if we consider 1) the incredible variety of organic molecules and their possible combinations, permutations, and interactions; 2) the incredible numbers of these molecules in the primitive oceans of the Earth; 3) the immense length of time available for the exploration of possibilities; 4) the steady energy cycles of day and night, the mixing action of wind, waves, and tides, the alternation of wet and dry conditions, the rhythmic cycles of Sun and Moon stirring the reaction vessel of Earth's oceans - under these circumstances the very improbable becomes the very likely. It is as if one person could buy all the tickets to the lottery. We envision the solar diurnal energy cycle as a kind of "CPR" or "artificial respiration", relentlessly applied to the apparently dead Earth by the Sun until the spark of life is kindled.
However, there is more order in this chemical system than random chance. This is not analogous to the situation of millions of monkeys typing on typewriters until, just by blind luck, one of them produces a Shakespearian sonnet. The key ingredient of life is the replication of molecular systems containing information. Without the ability to replicate, any advance in the information content of a system is lost, as no matter how sophisticated it may be, it will eventually be degraded and a new beginning will have to be made. With replication, however, information can be stored, retrieved, and through the evolutionary process, advanced. We really do not have to concern ourselves with anything other than the evolution of a replicating molecule which can also conserve information, since once this is achieved, this molecule will compete with its own copies for resources, and natural selection and evolution (even at the molecular level) will generate all the rest of the necessary complexity. In other words, we have in RNA/DNA a molecule which can learn. Because the 4x3 structure of carbon (and water) is projected into the 4x3 structure of RNA, the generation of this replicating molecule is not a matter of pure chance, and therefore neither is the generation of life. The intersection of the 4x3 universal fractal algorithm with a replicating molecular structure (RNA - DNA) means that life is common in the Universe - a definite (and perhaps calculable) probability of organic chemistry (plus water). This is why life arose on Earth (the "water planet") relatively quickly. (See: "Newton and Darwin: the Origin and Abundance of Life in the Cosmos".)
RNA links and catalyzes chemical systems which acquire, store, and degrade energy, systems which may well have evolved in response to the diurnal energy cycle. The emergent property of "metabolic" RNA is replication. The evolution of a replicative system from a metabolic mechanism for energy and enzyme transfer is helpful in terms of understanding the thermodynamic origin of, or reason for, reproduction: replication is the accidental byproduct of a 4x3 molecular structure originally involved in energy capture, or a related "metabolic" role, a chemical cycle driven, primed, or "pumped" by abiotic energy sources such as the Sun, Moon, and Earth's rotation. Energy capture, not replication per se, was the original "goal" or purpose of this complex chemical system; furthermore, its evolution was prompted by external, "mechanical", cyclic energy pulses from the Sun in which the darkness was as important as the light (in terms of posing an evolutionary or selective challenge - how to carry a functioning energy-capture system through the night - which is solved by memory-replication).
It is in row four that the "Chaos" model becomes especially compelling. The 4x3 form of RNA acts as a "strange attractor" or as a "fractal basin" for life, not just because it is a resonant 4x3 form, but because it is a resonant form capable of taking the next crucial step toward life - replication combined with system memory. Once molecular replication is achieved, the game is over, the lottery is won, the goal is reached. System memory in the form of replicating RNA survives the night to reap the rewards of a new day with its free energy inputs. A new "biological" dynamic takes over, featuring competition for limited resources and natural selection, and all the rest is evolutionary history. We place in cell 4 these cyclic, metabolic, replicating, prebiotic chemical systems; they will merge their separate abilities to become the coordinated elements of complex life forms in level five.
See also: "A Simpler Origin for Life" by Robert Shapiro. Scientific American, June, 2007, pages 46-53 (and on www.sciam.com). See also: "The Origin of Life" by James Trefil et. al. American Scientist Vol 97 No. 3, May-June 2009, pages 206 - 213.
The Biophysical Realm
Level 5: the Cell; DNA-RNA
(4 nucleotides code in triplets)
The Information Content of DNA
When we contemplate the molecular beginnings of life, we are initially at a loss to understand the motivation behind its chemical origins - what abiotic force could possibly drive a mixture of chemicals toward the organized, discreet, self-maintaining, self-reproducing, and self-interested (in the sense of survival) systems we call living entities? That DNA is a 4x3 fractal resonant system, evolved from a molecule (RNA) originally involved in an energy cycling system, which had the accidental, coincident, or emergent property of self-replication combined with chemical "memory", answers many of these questions; the rest are answered by evolution driven through natural selection in the frenzied competition for limited resources by a replicating population of (almost but not quite) identical molecular species.
In ourselves, we recognize the life force as our desire to breath, drink, and eat, driven by hunger, thirst, and the respiratory imperative; our desire to reproduce, driven by sexual desire; our desire to survive, driven by self-interest, fear, and pain; our desire for knowledge, driven by our need for food, mates, and survival, as well as an inherent and adaptive curiosity. At our highly evolved level of organization, we have no trouble identifying a number of forces which drive us individually and as a species to participate in the life process. It seems to me that these basic, instinctual, and physiological drives are nothing more than stage transformations of the same forces that drive the chemical system from its very beginning.
For example, DNA is at one and the same time a genetic information system, a metabolic (energy processing) machine, a replicating molecule, and a chemical memory system. When DNA replicates, it eats to do so - it must acquire from its environment the "food", or raw materials in the form of appropriate chemicals, to replicate its strands. But DNA's ability to bond to "appropriate chemicals" is also its combined information, memory, and perceptual system. In DNA these processes are inseparable, all belonging to the same act and molecule. All are driven by the forces of electrical attraction and repulsion between molecules, requiring also certain bonding strengths and geometric configurations. They simply scale up to our level of organization as the basic feelings, emotions, instincts, and drives which cause us to "want" to eat, reproduce, and perceive - since perception (information intake and processing) allows us to eat, survive, and reproduce more successfully. And because DNA is also a chemical memory system, our bodies remember or know how to perform all the basic functions of metabolism, respiration, reproduction, perception, growth, development, etc.
In humans, because of our higher level of organization, we have specialized organs which perform these functions separately - stomach, gonads, eyes, brain - so we think of them separately. But the bottom line is we must still eat to think or reproduce. There is less of a mystery here, in terms of the motivational inputs to chemical systems, than we are apt to believe at first. As for the complexity involved, staggering as it is, it is simply the product of evolution over eons of time and oceans of molecules. Similarly, we used to think the eye was too complex for evolution to produce; we know better now - even jellyfish and clams have them.
Once the replicating structure of DNA is achieved, it will compete with itself for the chemical resources to continue to reproduce. Competition within a species, with one's own kind, is the most powerful evolutionary force, and every time DNA replicates it produces a competitor whose needs are identical to its own. We can hardly imagine the ferocity of competition that must have taken place in the early oceans of Earth once this molecule began to replicate. The subsequent evolution of this molecule is simply the history of the most successful eater, reproducer, and defender of this adaptive behavior and information.
It is this competitive origin of the molecular system of life, natural selection operating within a domain of limited resources, that produces the anomalous "self interest" or "survival instinct" in even the most primitive form of life, and indeed, in the chemical systems which precede them.
I have no intention here of demoting the mystery or sanctity of life, only of understanding its mechanical elements. In my personal view, this mystery is not unique to our particular level of organization, but extends downward into the atoms of which we are composed, and upwards into the galaxies of which we are a part. Among many others, one of the enduring mysteries is the 4x3 organizational "metric" that propagates at every level from the Big Bang to the genome of life (see: "The Fractal Organization of Nature"). It is nevertheless true that there are mysteries associated with our own level of organization that are unique to the human experience. I will touch on these later.
As DNA is the molecular unit of replication, so the cell is the organizational unit of life, the "quantum entity" of biological systems. The cell is essentially a fortress or shelter erected against an inclement (and competitive) environment in which DNA can safely and efficiently go about its business of eating, reproducing, and storing the information necessary to do so. Within the protective confines of the cell wall, DNA can "get serious" about organizing its activities. In the same sense that "a man's home is his castle", the cell is the castle of DNA and the life process.
A house is not so different from a cell in its functional purpose. If our home is a farm, the analogy is better; if the barn with our symbiont livestock is attached to the house, the analogy is better still. At our organizational level (species) we as individuals are the functional analog of DNA and the house-office-barn is the cell which shelters us from the environment, but which also allows us to interact with it efficiently in ways we need and choose to. The knowledge and expertise we bring to our office, job, or profession is our personal information content which allows us to eat, reproduce, and survive. Communication with, and information gathering from the environment is of course crucial. In the modern house, we will want a mail box, a telephone, newspaper delivery, radio, TV, computer connected to the web, etc. There will be exits for wastes (sewer, garbage, ventilation) and entrances for supplies (water, electricity, antennas, gas, windows, doors, driveways), but even the most primitive shelter must have at minimum an entrance and exit. And like the cell, we are selective about who and what we let in and let out.
We do, of course, very much embody in our persons DNA going about its business of eating and reproducing as our information content allows. So it is really no wonder that our houses have evolved toward the ideal of the cell; indeed, in the contrast between the self-sufficient structures of the family farm and the interior cubicles of the modern high-rise business office, we see just the difference between the DNA of a free living cell and the DNA of a cell which has lost its independence to the higher organizational level of a corporate social organ of specialized function. It is partly from this loss of personal completeness and freedom that the angst of modern life arises. It reprises the difficulty of the evolutionary jump from single, free cells to the subjugation of individuality in the specialized functions of a higher level of organization, in our case the organism of society.
In cell 2 of row 5 we place the replication of the double-stranded DNA molecule; in cell 3 we place the cell nucleus and its genetic code. In cell 3 the four nucleotides of DNA code in triplets to produce the amino acids of proteins, the final molecular expression of the 4x3 organizational fractal - the simplest-sufficient or least-energy solution to the information code of biology and genetic heredity.
Here (cell 3) we should point out the role of symbionts in the functioning of the biological cell; this is an explicit expression of mutualism, symbiosis, and the "sharing of differences" which characterizes the third column. DNA itself is a collection of information and genes accumulated from many different sources. The role of symbionts in the formation and function of the cell - and in nature generally - has been pioneered and championed by Lynn Margulis and her coworkers. (See: Lynn Margulis and Dorion Sagan, 1997. Microcosmos: 4 Billion Years of Microbial Evolution. University of California Press, Berkeley.)
The biological cell is the first organizational level that we could describe as a complete "system", and the information content of this level is organizational or system information, having mostly to do with the orchestration of a multitude of integrated functions which all serve a single collective purpose: the feeding, defense, and reproduction of the cell. Here a variety of molecular subsystems are subjugated to the central control of the cell, in the form of nuclear DNA, which controls cellular function by means of RNA messengers, and whose genes turn on and off in response to chemical feedback (the so-called "epigenetic" system).
Yet, life is a cooperative venture, and within the cell there are at least 2 known symbionts, the chloroplasts and mitochondria, and others are suspected, such as the motile cilia. Chloroplasts and mitochondria (which produce and process energy for the cell) both have their own DNA and reproduce themselves independently within the cell. These are sub-cells which never agreed to relinquish their reproductive options to the larger organizational unit; but they, too, have made a separate peace and taken shelter within the larger group membrane. One advantage of this arrangement is that it relieves the central cell nucleus of the responsibility of reproducing these complex subsystems and carrying the information necessary to do so. They are in the role of "subcontractors" within the overall framework of the cell. It is clear that chloroplasts and mitochondria were originally free living cells, but they have found it more convenient to pursue their specialties within the shelter of a broader authority. Perhaps this allows them to sharpen their special skills, having less to worry about - much as the cook and mechanic on a ship can perform their duties better if they can leave the overall running of the ship to the captain. In turn, the cell and ship benefit from their enhanced, specialized functions.
In short, the cell is a miniature organism or society, complete with a central nuclear authority and specialized subsystems (both molecular and symbiotic) under its regulation. Could we doubt that such a system would eventually take the next organizational step and scale up to multicellular organisms? Life at the level of the single cell (bacteria, algae) is by far the most abundant, diverse, and resilient form of life on Earth. If humans ever establish colonies on other planets, we will (at least) take the bacteria with us, as symbionts which are necessary to digest our food, and which perform other vital system functions of which we may not yet be fully cognizant.
Level 6 - Organisms
(organs)
Multicellular organisms are the next stage beyond cellular systems. Here we find whole cells, rather than symbionts and chemical systems, subjugated to a central authority. In animals this central authority is vested in specialized cells, tissues, and organs such as the brain and hormone producing glands, which communicate with the entire body through the vascular and nervous systems, while in plants it seems to reside in hormones produced by the growing tips, which communicate with the rest of the plant through the sap. In either case, we find specialized functional cells, tissues and/or organs all synchronized and orchestrated into a harmonious whole.
It is a significant fact that it took nature much longer to evolve organisms from cells than it did to evolve living cells from chemicals. One explanation may be that evolution was working with a much smaller number of cells than of organic molecules. Another may be that it is very difficult to subjugate free-living cells to the central authority of an organism, especially to convince them to surrender their own reproductive prerogative to the specialized reproductive cells of an organism. In other words, evolution had to work against the resistance of the personal reproductive agendas of the cells, whereas in the earlier instance, molecular systems were not advanced enough to have "personal agendas" of any sort; hence the greater difficulty in this second stage of the evolutionary climb toward complexity. (Imagine the difficulty of organizing individual humans into a social system in which only a specialized few were allowed to reproduce sexually (and so achieve a potential immortality), while all the rest could only clone themselves temporarily. Clearly, this only works in the cells of an organism because they all carry exactly the same DNA.) To this day, mitochondria and chloroplasts have not relinquished their reproductive rights (they carry their own DNA), and we commonly suffer cancer. Finally, the orchestration of trillions of cells into a synchronized system of integrated specialized functions which grow, differentiate, reproduce, and pass through the different stages of life harmoniously together is clearly a task of extraordinary complexity. The large organisms that have populated the Earth for the last 3/4 billion years, plants as well as animals, are all marvels of systems engineering, systems information, and systems coordination, masterworks of the evolutionary process.
In the second position of row 6 we place cell division, and in the third, their differentiated product, the functionally specialized organ systems of the multicellular form. Of particular interest in this regard are the specialized organs of perception and their integration into a central processing center, the brain. The brain emerges as the central controlling authority, while perceptual organs function to gather information from the environment. Information can now be stored in the brain as memory, as well as in molecular codes.
Perceptual information is of central importance to all levels of life; in higher organisms it can be stored in memory as "software", rather than encoded into the "hardware" of the genome. Memory perishes with the death of the organism. Memory's utility is personal rather than general, and is a means of allowing an organism more flexibility in its behavior than is possible from its genome alone. Hence memory functions as a personal information set built up from the unique experience of the individual, which is in addition to its genetic information, the latter being approximately the same for all organisms of a given type. Memory is therefore a highly flexible and individually adaptive information set which adds greatly to the organism's chances of survival in a diverse and unpredictable environment. Memory itself (in the sense of the capacity for memory) becomes translated into genetic hardware through its contribution to the reproductive success of an organism. Individual memories can of course be transmitted as "software" to other organisms through the process of "learning", or by other cultural, social, or parental communication channels. The instinctual behavior of an ant colony, however, is an example of social "memory" transmitted in the form of genetics or molecular "hardware".
Level 7 - Species
(populations)
Species are defined by sexual reproduction and their closed genomes or exclusive gene pools. Cats and dogs cannot reproduce with each other, their gene pools are incompatible and exclude each other. The species level is our own level (Homo sapiens) of organization. This is the level of individuals, male and female, typically associated with the higher plants and animals, and of populations and social organization. While each individual contains a huge library of genetic information stored in the DNA of every cell, we see the emergence at this level of a new type of information carried in abstract as well as social form. Below level 7, information is carried in molecular form, in genes and hormones, in memory, and as electrical signals in nerve tissue.
At the species level we find information emerging in abstract form as signals, behavior, sounds, light, language - forms of information that have broken free of molecular carriers and are directed toward the specialized perceptual organs of sight and hearing. Smell, taste, and touch remain as more primitive (but nevertheless eminently useful) molecular and tactile information senses. While plants, lacking nervous systems, use only chemical communication, almost all animals, vertebrate and invertebrate, use some form of abstract communication. The songs and colors of insects and the flashing lights of fireflies are familiar, and invertebrates also use behavioral signals. Among the vertebrates, frogs croak, birds sing, bats and dolphins use sonar, snakes use heat, fish use light and electrical signals, the list is endless. The great whales are thought to be able to communicate by sound over enormous distances in the oceans.
Social organizations and communication allow personal memory to be shared and to persist from one generation to another. In humans, memory becomes encoded in writing and books stored in libraries, an information transformation which produces "social knowledge". In humans, the grammatical structure of language has raised the information content of speech to a new level of precision, abstraction, and beauty, and set in motion the explosion of human information systems which are the glory of our species, and which comprises a completely new, tangential evolutionary development (a new fractal iteration) within the "The Information Ladder".
The development of human abstract information systems has created an information explosion which is comparable only to that of organic chemistry in the microphysical realm, and genetics in the biological realm. Again, this information proliferation is the result of a lower energy threshold for interaction, abstract perceptual systems (sight, sound, language, writing, math, thought) vs molecular codes (genes). Will there be a another resonance of this information explosion in the astrophysical realm at a still lower energy threshold? (Super-cooled quantum computers? Machine intelligence?Psychic phenomena (ESP)? Deep meditation? Communication with aliens?)
Most higher animal species are divided into male and female individuals, an arrangement necessary to maintain the diversity of their otherwise closed gene pools. We also find social behavior at this level, which presages the emergence of a higher organization level, the social organism, fully developed among the invertebrates (corals, termites, ants, bees) and (thankfully) developed to a considerably lesser degree of perfection in the political, religious, economic, and military organizations of humans. There is strength and safety in numbers, and efficiency and opportunity in specialization, provided all is prudently organized.
In cell two of level 7 we place sexual reproduction, male and female, and in cell three social organization, in which a population of individuals (a subset of the whole species), is organized into groups with specialized functions all coordinated by a central authority. In humans, this authority is formalized as government, a mixture of empowered individuals and abstract organizational codes, system rules, or laws.
The purpose of social organization is to enhance individual survival and reproductive success through coordinated group action and the efficiency of functional specialization. Human society can advance rapidly through the contributions of its most talented individuals because the society can capture their personal experience, knowledge, and insights in writing, and incorporate their personal expertise into social memory, knowledge, and function. An additional benefit of social life is personal access to social products which no individual could produce alone. In humans this "corporate dividend" has come to include the technology and cities of the current era.
Level 8 - Gaia
(ecosystems)
The final biophysical level of information/organization is that of the ecosystem of the entire planet, approximately the holistic concept of "Gaia" advanced by J. E. Lovelock. At this organizational level, species are but the units in a broader classification of herbivores, predators, parasites, producers (plants), consumers (animals), scavengers and detritivores (agents of decay), etc. Species exist in aquatic and terrestrial habitats, distributed over the surface of the globe in response to climatic cycles, continental drift, and evolutionary pressures over geologic time (biogeography). The Gaian hypothesis is simply that life modifies the physical environment of Earth on a global scale in ways that are conducive to life's continuation - the Earth has been and continues to be "terraformed" by life to create a more suitable and stable habitat for itself.
These effects are most notable in the chemical composition of the atmosphere, oceans, and soils of the planet, and the temperature, rainfall, and perhaps even the climatic cycles of the Earth. The fact that the chemical composition of Earth's atmosphere, its temperature, and even the chemistry and salinity of the oceans, has remained stable over geologic time is evidence of the homeostatic, self-serving activity of Gaia.
The grand symbiosis between plants and animals with respect to the atmospheric gases oxygen, carbon dioxide, and the availability of nitrogen are part of this interplay between the physical and biotic environment, as are biogeochemical cycles involving the formation of petroleum, gas, and coal deposits, limestone and salt beds, etc. The positive feedback between plants and rainfall, the greenhouse effect on climate, etc., are other examples of the linkage between living systems and the abiotic environment. Genetic diversity, heritability, and the competition for limited resources continues to generate new species over evolutionary time which exploit the environment in new and/or more efficient ways, and each new species is itself a new resource for some other species. Natural checks and balances, such as predators, parasites, and disease organisms, competition, the harshness, limitations, diversity, and unpredictability of the environment, ensure that no one species oversteps the boundaries of a balanced system for long. Symbiotic relationships are favored over exploitive relationships in the long run. Thus the extinction of the horse by early hunters in North America may be contrasted with its domestication in Asia, to the great benefit in the latter continent of both species. Similarly, agriculture as a symbiotic relationship is greatly favored over hunting and gathering as a method of obtaining food.
The glacial cycles of the recent geologic history of Earth show that extreme climatic shifts can occur even within the context of a fully formed, abundant, and diverse biota, and yet the natural system contains some type of internal buffer or negative feedback that causes the glaciers to retreat more rapidly than they advanced. We would have to suspect that these cycles have both a biotic and an abiotic cause (such as the Milankovitch orbital cycles of planet Earth, with perhaps an additional input from continental drift).
The Biological Role of
Humanity - From Gaia's Perspective
(why we are here and what we should do)
But for the peculiar appearance of Man amongst the myriad species of Gaian life forms, this would be the end of our story - the planet Earth, overflowing with life but isolated in the vastness of space, a magnificent experiment in the building of biological information systems of incredible diversity and complexity, all doomed to die with the inevitable exhaustion of our Sun. But Gaia has not gone to all this trouble for nothing, for in Homo sapiens she has planned her escape from the dying Sun, or catastrophic asteroid impact. Human Space Travelers will carry Gaian life into the galaxy in every direction as far as possible, spreading like bacteria upon an agar plate until we encounter occupied ground and the counter-thrust of a similarly spreading but alien life form.
Gaia is in her reproductive phase, and we are her seeds, pawns of her reproductive purposes. She has created us as dispersal agents for herself. The vision of Noah's ark is a vision of the future, not the past. This simple fact explains all of human evolution, every capacity of Man, and all the abundance of resources which Gaia has stored over the eons for our use, so that we may accomplish her great reproductive goal. Like every other life form, Gaia has her season of flowering, reproduction, and dispersal to new territory. This is the special biological meaning of Man and the special significance of the time in which we live. The new millennium will be the age of space travel and discovery, just as the last has been the age of ocean travel and discovery upon the Earth.
We think the space program is our program; it is not. Humanity and the space program have both been produced by Gaia.
We have already entered upon the threshold of step 9, the exploration and colonization of the Solar System. This will be the work of the 21st century and beyond. We cannot tell when the next stages will occur - they depend upon too many unknowns at our current stage of development. But we see where the information pathway leads, and our art - in the form of science fiction - is already at the stage of galactic exploration and interaction with aliens, who are understandably busy doing just what we will be doing - grabbing as much suitable territory for their planet's life form as possible. We can only hope these future interactions will be symbiotic. Hence intelligent life is a way for planets to interact across the vastness of space, just as atomic nuclei interact at great relative distances through the electron shell, and cells communicate with each other through the vascular systems of organisms, which in turn communicate through space via the signals and sounds of innumerable species.
Gaia is the first level in the table of organization which is above our own level, and in level 8 we find forces and purposes which are greater than our own, which we may study and come to understand but never control, and to which we are in many ways subservient. It is obvious, for example, that at the planetary level we will never control the forces of the Solar System - the evolution of the Sun, or the orbits of the moon or other planets. What is perhaps not so obvious is that this same lack of control exists at the level of Gaia, the natural forces of the Earth herself. We will never control plate tectonics, volcanism, or earthquakes, nor, it is likely, the weather or the cycles of climate. It furthermore appears that we are not even in control of our own destiny, in the sense that we are but the agents of Gaia's reproductive intent. Hopefully, what is good for Gaia is also good for us (we must accept this as a basic truth, just as every cell in our body must accept the equivalent premise), but we are used to thinking of ourselves as the masters of the Earth, not the pawns of Her reproductive schemes.
It is certainly possible that Gaia will be significantly weakened by her reproductive effort; that humans will be forced off the planet to escape their own aggression, exploitation, pollution, and overpopulation. In this case the expulsion from the Garden of Eden, like the story of Noah's Ark, will also come to be seen as a vision of the future rather than the past.
In a purely biological sense, we have found the answer to the great question of existence - why are we here, who are we, and what should we do? We are the reproductive seeds of Gaia and we exist to colonize the galaxy with her life forms. From the perspective of Gaia, that is why humanity has been brought forth upon the Earth. Therefore we should do everything we can to maximize our cooperative conquest of space - including stopping fighting among ourselves and protecting our Earth, which is the only and best life-support system we will have in this galaxy for a very long time.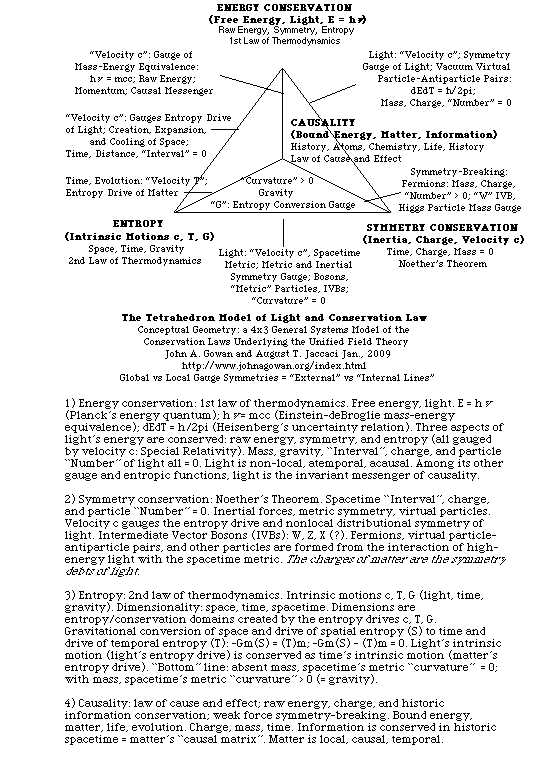{kind=link}